

Esponjas reproduzem-se clonalmente (assexuadamente) por fragmentação, brotamento e pela formação de págulos de resistência, chamados de gêmulas. A fragmentação é principalmente o resultado de danos oriundos de correntes e ondas e, talvez de danos causados por carnívoros pastadores, os fragmentos desalojados dependem de sua capacidade de reestruturação por regeneração. O fragmento logo se prende ao substrato e se reorganiza em uma esponja funcional. O brotamento é incomum, mas acontece em algumas esponjas. Certas espécies de Tethya produzem brotos penduculados. Espécies de Oscarella e Aplysilla foram reportadas como produzindo papilas que se auto-amputam e crescem como novas esponjas.
Muitas esponjas de água doce e algumas espécies marinhas produzem centenas a milhares de gêmulas semelhantes a esporos, tipicamente no outono do ano. As gêmulas de outono das espécies de água doce podem entra em diapausa, um estado próximo da suspensão metabólica, requerendo, assim, um período de temperatura muito fria antes que sejam novamente ativadas, germinem e diferenciem-se em uma nova esponja, normalmente na primavera. Enquanto a gêmula estiver em diapausa, é resistente aos extremos ambientais de temperatura, salinidade e dessecação.
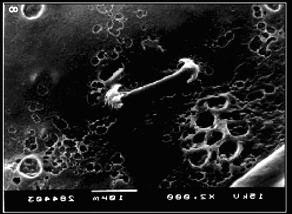
Gêmulas são produzidas no meso-hilo de uma esponja agonizante ao redor de um agrupamento de arqueócitos carregados de nutrientes. Espongiócitos secretam um envoltório de espongina ao redor da massa celular. O envoltório também pode conter espículas secretadas por esclerócitos. O envoltório inclui completamente a massa de células exceto em um pólo em que a abertura, a micrópila.
A gêmula completa consiste e um envoltório que inclui seus arqueócitos, cada um dos quais logo se tornaram esféricos, assemelhando-se a uma célula embrionária e, então, chamados de tesócitos. Durante a primavera a gêmula “eclode”, os tesócitos periféricos se diferenciam em uma pinacoderme que migra para o exterior, como um abola de chiclete, pela micrópila. Essa “bolha” de pinacoderme estabelece contato e adere ao substrato. Depois, os tesócitos mais profundos extravasam pela micrópila para dentro da bolha e estabelecem, após a diferenciação, o interior da esponja jovem.
Reprodução Sexuada
Esponjas com poucas exceções são hermafroditas. No momento apropriado, espermatozóides são gerados e liberados de uma esponja, transportados pelas correntes d’água a outras nas quais a fertilização acontece interiormente. Algumas espécies (como Cliona) são ovíparas e liberam zigotos na água, onde eles completam seu desenvolvimento. A maioria das esponjas é vivípara, retendo os zigotos no corpo parental e liberando larvas (as vezes chamado de larviparidade). Embriões e larvas são lecitotróficas.
As esponjas são consideradas como não possuindo órgãos genitais (gônadas) e células germinativas acontecem tanto em conjuntos simples (espermatozóides) ou individualmente (óvulos) ao longo do meso-hilo. Espermatozóides surgem de coanócitos ou câmaras coanociticas completas que se aprofundam no meso-hilo e são envolvidas em uma fina parede celular para formar um cisto espermático. Óvulos surgem de arqueócitos ou coanócitos desdiferenciados (em algumas esponjas calcárias). Cada óvulo geralmente acumula seu vitelo por fagocitose de células nutritivas adjacentes. O óvulo e células nutritivas podem ser incluídos juntos em um folículo de células encistadas. Como o sistema aqüífero supre todas as partes do corpo igualmente, as células germinativas são também amplamente distribuídas ao longo do meso-hilo do corpo, mas sempre dentro da distância de difusão de um canal ou câmara.
Durante a liberação, os espermatozóides rompem a parede do cisto espermático, entram nos canais exalantes (ou átrios) e são liberados pelos ósculos. Certas espécies tropicais, conhecidas dos mergulhadores como esponjas de fumaça, repentinamente liberam esperma em nuvens brancas a partir de seus ósculos. Tal liberação súbita de esperma pode ser típica para a maioria das esponjas
 Quando o esperma liberado deriva e contata outra esponja, eles são passados para seu sistema aqüífero pelo fluxo de água inalante. Uma vez no sistema aqüífero, o espermatozóide é transportado para a coanoderme ou câmara coanocítica e é fagocitado, mas não digerido, por um coanócito. Este, então, perde seu flagelo e colarinho, se torna amebóide e transporta a cabeça do espermatozóide (núcleo) para o óvulo. O coanócito amebóide transformado é chamado de célula transportadora. Depois que a célula transportadora alcançar um óvulo no meso-hilo próximo, ou ela transfere o núcleo do espermatozóide ao óvulo ou a célula transportadora e núcleo do espermatozóide são juntos fagocitados pelo óvulo. De qualquer maneira, a fertilização acontece internamente no “ovário” da esponja.
Quando o esperma liberado deriva e contata outra esponja, eles são passados para seu sistema aqüífero pelo fluxo de água inalante. Uma vez no sistema aqüífero, o espermatozóide é transportado para a coanoderme ou câmara coanocítica e é fagocitado, mas não digerido, por um coanócito. Este, então, perde seu flagelo e colarinho, se torna amebóide e transporta a cabeça do espermatozóide (núcleo) para o óvulo. O coanócito amebóide transformado é chamado de célula transportadora. Depois que a célula transportadora alcançar um óvulo no meso-hilo próximo, ou ela transfere o núcleo do espermatozóide ao óvulo ou a célula transportadora e núcleo do espermatozóide são juntos fagocitados pelo óvulo. De qualquer maneira, a fertilização acontece internamente no “ovário” da esponja.
O espermatozóide da maioria das esponjas não possui um acrossomo, a estrutura responsável em penetrar a membrana da célula-ovo durante a fertilização na maioria dos outros animais. Um acrossomo provavelmente é desnecessário porque o núcleo do espermatozóide entra no ovo por fagocitose. O acrossomo espermatozóide ocorre em Oscarella lobularis, sugerindo que ela possui meios convencionais de fertilização do ovulo, mas os detalhes reprodutivos dessa espécie são desconhecidos.
O zigoto sofre clivagem holoblástica em blastômeros de tamanhos iguais. O padrão de células que resulta da clivagem, porém, varia entre as espécies de esponjas. As larvas que se desenvolvem de embriões também são diversas e descritas com os nomes de larvas celoblástula, anfiblástula, parenquimela e triquimela.
Referências Bibliograficas:
Ruppert, Edward E.
Zoologia dos invertebrados: uma abordagem funcional - evolutiva / Edward E. Ruppert; Richard S. Fox; Robert D. Barnes; [revisão cientificaAntonio Carlos Marques, cordenador e revisor da tradução]. São Paulo; Roca, 2005



Nenhum comentário:
Postar um comentário